
Type Species: Ceratosaurus nasicornis
Classification: Dinosauria - Saurischia – Theropoda – Neotheropoda – Ceratosauria – Neoceratosauria – Ceratosauridae
Time Period: Late Jurassic
Location: North America
Diet: Carnivore
The ‘horned lizard’ Ceratosaurus is one of the main theropods that prowled North America during the Late Jurassic. This dinosaur was discovered early in paleontology, and so its name was given to one of the major theropod subgroups, the Ceratosauria, a sister taxon to the ‘stiff-tailed’ tetanurans. Members of Ceratosauria had robust skulls with increased ornamentation or height and a shortening of the arms. Ceratosaurs are identified as theropods closer in relation to Ceratosaurus than to birds; indeed, it was the tetanuran line that eventually diversified into modern birds. Though Ceratosauria diverged from the rest of theropoda – and its sister taxon Tetanurae – in the Early Jurassic, most ceratosaurs were from the Late Jurassic (with a few identified in the Cretaceous).

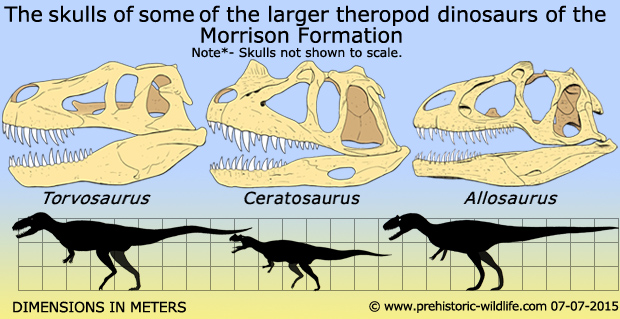
Three nearly complete Ceratosaurus specimens have been uncovered (along with a host of fragmentary remains) in the North American Morrison Formation. Though possible remains have also been found in Africa, South America, and Europe, the identification of these fossils as belonging to Ceratosaurus is dubious; thus, while some would like to push Ceratosaurus’ domain southward and eastward from North America, more research is needed before this can be done in good faith. What is not in doubt is that Ceratosaurus was a ‘staple theropod’ of the Morrison Formation. Radiometric dating puts the age of the Morrison Formation and its associated fossil-beds to between 156 and 146 million years ago, safely nestled in the Late Jurassic and including the late Oxfordian stage, the Kimmeridgian stage, and the early Tithonian stage of the period. The Morrison Formation was a semiarid environment with distinct wet and dry seasons. The Morrison Basin stretched from New Mexico to Alberta and Saskatchewan, and was formed when the precursors to the Front Range of the Rocky Mountains started pushing up to the west. The deposits from their east-facing drainage basins were carried by streams and rivers and deposited in swampy lowlands, lakes, river channels, and floodplains. These environments were dominated by the herbivorous sauropods, though it was also home to the plated Stegosaurus and early ornithopods such as Camptosaurus. The top predator of the area was Allosaurus, which accounts for up to seventy-five percent of theropod specimens. Non-dinosaurian organisms included ray-finned fishes, frogs, salamanders, turtles, lizards, terrestrial and aquatic crocodylomorphs, and several species of pterosaurs. Fossilized flora of the Morrison Formation includes green algae, fungi, mosses, horsetails, cycads, ginkgoes, and conifers. The Morrison Formation was a wide tract of territory with sub-environments that included river-lined forests of tree ferns and ferns (called ‘gallery forests’); swampy ‘backwaters’; lakeside ‘resorts’; and fern savannas with occasional trees such as the Araucaria-like conifer Brachyphyllum.

Ceratosaurus reached between seventeen and twenty-three feet in length and weighed up to a ton. Researchers have called it the ‘dragon’ of the theropods, because it had a striking resemblance to dragons of lore: it had a short, ridge-like horn on the tip of its snout, horny ridges near its eyes, and a jagged crest of osteoderms running down its neck. Fossil remains of the nasal horn include only the horn core; in life, the core would’ve been supported by a keratinous sheath. While the base of the nasal horn was smooth, its upper two-thirds were wrinkled and lined with grooves that contained blood vessels in life. In the holotype specimen, the horn core was five inches long and three-quarters of an inch wide at its base, but it narrowed to only half an inch wide further up the snout. It was just under three inches in height. In another specimen, it was longer and lower in the skull; this may be an episode of sexual dimorphism (perhaps males had larger horns than females?). These horns would’ve been larger in life due to the keratinous sheath. Though scientists used to believe the horn was used as a weapon while hunting prey, more studies have shown it was too small and weak to be used as a weapon. Though a few researchers believe it was used in interspecies combat between males – perhaps to establish territory or to determine mating rights – most scientists believe its function was strictly for display. When the nasal horn fused in maturity, it may have signaled the male’s readiness to mate. In this scenario, the nasal horn was primarily a function for males. It may have been richly colored in life. Ceratosaurus also had smaller, semicircular, bony ridges in front of each eye, similar to those of Allosaurus. These ridges were formed by the lacrimal bones. In juveniles, all three horns were smaller than in adults, and the two halves of the nasal horn didn’t fuse until later in life. Ceratosaurus’ further ornamentation included small, elongated, and irregularly formed osteoderms (skin bones) that ran along the midline of its body, likely forming a continuous row that extended from the base of the skull to most of the tail.
Ceratosaurus’ arms were short but strong and capped with four fingers on the hands (a primitive condition, as more ‘developed’ theropods would be three-fingered or even two-fingered). Its powerful rear limbs made it at a capable runner, and its skull was two feet long. It had massive fanged teeth set in deep jaws; the maxillary bones of the upper jaw were lined with fifteen blade-like, serrated teeth on each side. The first eight of these teeth were long and robust, but from the ninth tooth onward they gradually decreased in size. Ceratosaurus’ skull was unique in that it had cranial kenesis, in which the bones were so loosely connected that the skull bones could move side-to-side when eating; this enabled Ceratosaurus to swallow large pieces of meat. Ceratosaurus’ hunting methods have been a significant debate, largely because it shared its environment with larger theropods such as Allosaurus and Torvosaurus. Because it shared its environment with ‘bigger and badder’ theropods, scientists believe these predators practiced niche partitioning, in which they didn’t directly compete for food. One suggestion is that Ceratosaurus, being the smaller theropod of the Morrison ‘big-wigs’, hunted smaller ornithopods and stegosaurs whereas Allosaurus and Torvosaurus hunted larger herbivores such as the sauropods. Other scientists have proposed two more ‘developed’ theories regarding how niche partitioning may have played out between these theropods, and these theories have been promoted, embraced, and fought over; the first categorizes Ceratosaurus as a terrestrial hunter, and the second categorizes Ceratosaurus as an aquatic hunter who no doubt scavenged large dinosaur carcasses when the opportunity arose.
In 1998 paleontologist Donald Henderson suggested that Ceratosaurus lived alongside two separate potential species of Allosaurus. The first Allosaurus ‘morph’ had a shortened snout, a high and wide skull, and short, backwards-angled teeth; the second Allosaurus ‘morph’ had a longer snout, lower skull, and long, vertical teeth. Henderson attributed these morphological differences to variation in feeding strategies, so that the short-snouted Allosaurus occupied a different niche than both the long-snouted Allosaurus as well as Ceratosaurus. The short-snouted Allosaurus would’ve had reduced movement when biting, resulting in increased bite force, similar to what we see in cats. The long-snouted Allosaurus and Ceratosaurus would’ve used their fang-like teeth to deliver quick, slashing bites; their narrower skulls resulted in a generally weaker bite force. Because the long-snouted Allosaurus and Ceratosaurus would’ve had similar feeding strategies, Henderson suggested they would’ve been in direct competition in their environment; because only one species could reign supreme, the ‘loser’ would have to find elsewhere to hunt. Henderson argues that this is precisely what happened, and the larger long-snouted Allosaurus was triumphant. He shows how Ceratosaurus remains are rare in environments that include the long-snouted Allosaurus, but Ceratosaurus and the short-snouted Allosaurus – both employing different feeding strategies – could be found together. While this theory is intriguing, some oppose it by arguing that the two Allosaurus ‘morphologies’ may not be as ‘cut and dry’ as Henderson thinks. Bolstering skeptics’ cause, a 2010 study suggested that short-snouted Allosaurus were actually cases of individual variation rather than a separate species.

Henderson’ theory assumes that Ceratosaurus was a terrestrial predator, but a 2004 study by Robert Bakker and Gary Bir went another direction: “What if Ceratosaurus specialized in aquatic prey, feeding on fish and crocodiles and turtles?” They pointed out how shed teeth from fifty different localities in the Morrison Formation belong to both Ceratosaurus and megalosaurs tend to show up in habitats in and near water sources such as wet floodplains, riverine locations, lake margins, and swamps. While allosaurids were common in both terrestrial and aquatic environments, Ceratosaurus was predominantly found in the latter. Bakker and Bir argued that Ceratosaurus and megalosaurs preferred to hunt near and in bodies of water (though they no doubt fed on scavenged dinosaur carcasses; Ceratosaurus tooth marks have been found on sauropod bones that it likely scavenged). They pointed out how Ceratosaurus and megalosaurs in general had long, low, and flexible bodies. Compared to other Morrison theropods, Ceratosaurus had taller neural spines on the foremost tail vertebrae, which were vertical rather than inclined toward the back; these, when combined with the deep chevron bones on the underside of the tail, indicate a deep, ‘crocodile-like’ tail that may have been designed for swimming. Allosaurids, in contrast, had shorter, taller, and stiffer bodies with longer legs; Allosaurus and its ilk were designed for rapid running in open terrain and preying upon larger sauropods and stegosaurs. The Morrison Formation, resplendent with swamp-lands and lakes and river valleys, would’ve been a prime environment for a primarily aquatic hunter such as Ceratosaurus. Perhaps, if we were to step back in time and walk along the shore of a Jurassic river, we would see a Ceratosaurus sculling its way through the current on the hunt for crocodilians and fish.
No comments:
Post a Comment